Da bi se sveobuhvatno moglo govoriti o hemoglobinu (Hb), korisno je prvo se baviti mioglobinom (Mb) koji je vrlo sličan hemoglobinu, ali je mnogo jednostavniji. Među hemoglobinom i mioglobinom postoje strogi odnosi srodstva: oba su konjugirana proteina, a njihova protetska skupina (ne-proteinski dio) je heme skupina.
Mioglobin je globularni protein koji se sastoji od jednog lanca od oko stotinu i pedeset aminokiselina (ovisno o organizmu), a njegova molekularna težina je oko 18 Kd.
Kao što je spomenuto, opremljen je s heme skupinom koja je umetnuta u hidrofobni (ili lipofilni) dio proteina, koji se sastoji od nabora koji se mogu pripisati α-helix strukturama vlaknastih proteina.
Mioglobin se sastoji uglavnom od segmenata α-spirala, prisutnih u broju osam i sastoji se, gotovo isključivo, od nepolarnih ostataka (leucin, valin, metionin i fenilalanin) dok su polarni ostaci praktički odsutni (asparaginska kiselina, glutaminska kiselina, lizin i arginin); jedini polarni ostaci su dva histidina, koji igraju temeljnu ulogu u napadu kisika na heme skupinu.
Heme skupina je kromoforska skupina (apsorbira se u vidljivom) i funkcionalna je skupina mioglobina.
Vidi također: glikirani hemoglobin - hemoglobin u urinu
Malo kemije
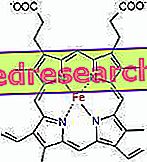
Veza između protoporfirina i željeza tipična je veza takozvanih koordinacijskih spojeva koji su kemijski spojevi u kojima središnji atom (ili ion) formira veze s drugim kemijskim vrstama u broju koji je veći od njegovog oksidacijskog broja (električni naboj). U slučaju heme, te su veze reverzibilne i slabe.
Koordinacijski broj (broj koordinacijskih veza) željeza je šest: oko željeza može biti šest molekula koje dijele elektrone koji se vežu.
Za formiranje koordinacijskog spoja potrebno je dvije orbitale s ispravnom orijentacijom: osoba može "kupiti" elektrone, a druga ih može donirati
U hemu, željezo tvori četiri ravninske veze s četiri atoma dušika koji su u središtu pralnog porfirinskog prstena i peta veza s proksimalnim histidinskim dušikom; željezo ima šestu slobodnu koordinacijsku vezu i može se vezati za kisik.
Kada je željezo u obliku slobodnog iona, njegove orbite d- tipa imaju istu energiju; u mioglobinu, željezni ion je vezan za protoporfirin i histidin: te vrste magnetski remete orbitale željeza; opseg poremećaja će biti različit za različite orbitale u skladu s njihovom prostornom orijentacijom i onom ometajućih vrsta. Budući da se ukupna energija orbitala mora vratiti na konstantu, perturbacija uzrokuje energetsko razdvajanje između različitih orbitala: energija dobivena nekim orbitalima, ekvivalentna je energiji koju su izgubili drugi.
Ako razdvajanje koje se događa između orbitala nije jako veliko, poželjan je elektronički raspored visokih spina: elektroni koji se vežu pokušavaju se paralelno okretati u nekoliko mogućih pod-razina (maksimalna množina); ako je, s druge strane, perturbacija vrlo jaka i postoji velika razdvojenost između orbitala, prikladnije je upariti elektrone za vezanje u orbitale niže energije (niski spin).
Kada se željezo veže na kisik, molekula poprima nisku raspodjelu spina, dok kada željezo ima šestu slobodnu koordinacijsku vezu, molekula ima visoki spin.
Zahvaljujući toj razlici u spinu, kroz spektralnu analizu mioglobina, možemo razumjeti postoji li vezani kisik (MbO2) ili ne (Mb).
Mioglobin je protein karakterističan za mišiće (ali ne samo u mišićima).
Mioglobin se ekstrahira iz kita u kojem je prisutan u velikim količinama, a zatim pročišćava.
Cetaceani imaju disanje kao ljudska bića: moraju imati pluća apsorbiraju zrak kroz respiratorni proces; kit-spermatozoid mora nositi što je više moguće kisika u mišićima koji mogu nakupiti kisik tako da ga vežu na prisutni mioglobin; Kiseonik se zatim polagano oslobađa kada se kitovi uranjaju jer mu je za metabolizam potreban kisik: što je veća količina kisika koju kit može moći apsorbirati i više kisika je dostupno tijekom ronjenja.

Da bi meso bilo manje ili više crveno je sadržaj hemoproteina (to je heme koje čini meso crvenom).
Hemoglobin ima mnoge strukturne analogije s mioglobinom i sposoban je vezati molekularni kisik reverzibilno; ali, iako je mioglobin ograničen na mišiće i periferna tkiva općenito, hemoglobin se nalazi u eritrocitima ili crvenim krvnim stanicama (to su pseudo-stanice, tj. one nisu stvarne stanice) koje čine 40% krvi.
Hemoglobin je tetrametar, to jest, sastoji se od četiri polipeptidna lanca, svaki s heme skupinom i dva po dva (u ljudskom biću postoje dva alfa lanca i dva beta lanca).
Glavna funkcija hemoglobina je transport kisika ; Druga funkcija krvi u kojoj je uključen hemoglobin je transport tvari u tkiva.
Na putu od pluća (bogat kisikom) do tkiva, hemoglobin prenosi kisik (u isto vrijeme i druge tvari dopiru do tkiva) dok na obrnutom putu nosi sa sobom otpad prikupljen iz tkiva, posebno ugljični dioksid koji nastaje u metabolizmu.
U razvoju čovjeka postoje geni koji se izražavaju samo određeno vrijeme; zbog toga imamo različite hemoglobine: fetalni, embrionalni, odraslog muškarca.
Lanci koji sačinjavaju ove različite hemoglobine imaju različite strukture, ali sa sličnostima funkcija koju obavljaju manje ili više je ista.
Objašnjenje prisutnosti nekoliko različitih lanaca je kako slijedi: tijekom evolucijskog procesa organizama, hemoglobin se također razvio specijalizirajući se za transport kisika iz područja koja su bogata u deficitarnim područjima. Na početku evolucijskog lanca hemoglobin je prenosio kisik u malim organizmima; tijekom evolucije organizmi su dostigli veće dimenzije, stoga se hemoglobin promijenio kako bi mogao transportirati kisik u područja dalje od mjesta gdje je bio bogat; da bi se to postiglo, nove strukture lanaca koje čine hemoglobin kodificirane su tijekom evolucijskog procesa.
Mioglobin veže kisik čak i kod skromnih pritisaka; u perifernim tkivima postoji pritisak (PO2) od oko 30 mmHg: na ovom pritisku mioglobin ne oslobađa kisik, pa bi bio neučinkovit kao nosač kisika. Hemoglobin, s druge strane, ima više elastičnog ponašanja: veže kisik pri visokim tlakovima i oslobađa ga kada pad pritiska.
Kada je protein funkcionalno aktivan, može malo promijeniti svoj oblik; na primjer, oksigenirani mioglobin ima drugačiji oblik od neogljičnog mioglobina i ta mutacija ne utječe na susjedne.
Diskurs je drugačiji u slučaju pridruženih proteina kao što je hemoglobin: kada je lanac oksidiran, on se inducira da promijeni svoj oblik, ali ova modifikacija je trodimenzionalna pa su tako i drugi lanci tetrametra. Činjenica da su lanci povezani jedan s drugim dovodi do mišljenja da modifikacija jednog utječe na druge susjedne, čak i ako je u različitoj mjeri; kada je lanac oksigeniran, drugi lanci tetrametera imaju "manje neprijateljski stav" prema kisiku: poteškoća s kojom se lanac oksigenata smanjuje kako se lanci uz njega oksigeniraju sami. Isti argument vrijedi i za deoksigenaciju.
Kvartarna struktura deoksihemoglobina naziva se T (napeta) forma, dok se oksihemoglobin naziva R oblik (oslobođen); u rastegnutom stanju postoji niz prilično jakih elektrostatskih interakcija između kiselinskih aminokiselina i bazičnih aminokiselina koje dovode do krute strukture deoksihemoglobina (zbog toga je "napeti oblik"), dok je kada je vezan kisik, entitet interakcije se smanjuju (to je razlog zašto "oslobođeni oblik"). Nadalje, u odsutnosti kisika, histidinski naboj (vidi strukturu) stabiliziran je suprotnim nabojem asparaginske kiseline, dok u prisutnosti kisika postoji tendencija da protein gubi proton; sve to znači da je oksigenirani hemoglobin jača kiselina od deoksigenirane hemoglobije: bohrov efekt .
Ovisno o pH, heme grupa se više ili manje lako veže na kisik: u kiselom okolišu hemoglobin lakše oslobađa kisik (napeti oblik je stabilan), dok je u bazičnoj sredini veza s kisikom jači.

Svaki hemoglobin oslobađa 0.7 protona po dolaznom molu kisika (O2).
Bohrov efekt omogućuje hemoglobinu da poboljša svoju sposobnost prenošenja kisika.
Hemoglobin koji putovanje od pluća do tkiva mora uravnotežiti prema tlaku, pH i temperaturi.
Vidimo učinak temperature .
Temperatura u plućnim alveolama je oko 1-1, 5 ° C niža od vanjske temperature, dok je u mišićima temperatura oko 36, 5-37 ° C; Kako se temperatura povećava, faktor zasićenja se smanjuje (s istim tlakom): to se događa zbog povećanja kinetičke energije i favoriziranja disocijacije.

Postoje i drugi čimbenici koji mogu utjecati na sposobnost hemoglobina da se veže na kisik, a jedan od njih je koncentracija 2, 3-bisfosfoglicerata .
2, 3-bisfosfoglicerat je metaboličko sredstvo prisutno u eritrocitima u koncentraciji od 4-5 mM (ni u kojem drugom dijelu tijela nije prisutno u tako visokoj koncentraciji).
Pri fiziološkom pH, 2, 3-bisfosfoglicerat se deprotonira i na njemu ima pet negativnih naboja; ona ide u klin između dva beta lanca hemoglobina jer ti lanci imaju visoku koncentraciju pozitivnih naboja. Elektrostatske interakcije između beta lanaca i bisfosfogliceratnih lanaca daju određenu krutost sustavu: dobiva se napeta struktura koja ima mali afinitet za kisik; tijekom oksigenacije, tada se izlučuje 2, 3-bisfosfoglicerat.
U eritrocitima postoji poseban aparat koji pretvara 1, 3-bisfosfoglicerat (proizveden metabolizmom) u 2, 3-bisfosfoglicerat tako da dosegne koncentraciju od 4-5 mM i stoga je hemoglobin u stanju razmijeniti kisik u tkivima.
Hemoglobin koji stiže u tkivo je u oslobođenom stanju (vezan za kisik), ali u blizini tkiva je karboksiliran i prelazi u rastegnuto stanje: protein u ovom stanju ima nižu tendenciju vezanja s kisikom, u usporedbi s oslobođenim stanjem, stoga hemoglobin oslobađa kisik u tkivo; nadalje, zbog reakcije između vode i ugljičnog dioksida dolazi do stvaranja iona H +, dakle daljnjeg kisika zbog efekta bohra.
Ugljični dioksid difundira u eritrocit koji prolazi kroz membranu plazme; s obzirom da eritrociti čine oko 40% krvi, treba očekivati da samo 40% ugljičnog dioksida koji difundira iz tkiva ulazi u njih, u stvarnosti 90% ugljičnog dioksida ulazi u crvene stanice jer sadrže enzim koji se pretvara ugljični dioksid u ugljičnoj kiselini, slijedi da je stacionarna koncentracija ugljičnog dioksida u eritrocitima niska i stoga je brzina ulaska visoka.

Još jedna pojava koja se javlja kada eritrocit dođe do tkiva je sljedeći: gradijentom HCO3- (derivat ugljičnog dioksida) izlazi iz eritrocita i, da bi uravnotežili izlaz negativnog naboja, imamo ulazak klorida koji određuje povećanje osmotskog tlaka: da bi se uravnotežila ova varijacija dolazi i do ulaska vode koja uzrokuje oticanje eritrocita (efekt HAMBURGER). Suprotan fenomen javlja se kada eritrocit dosegne plućne alveole: dolazi do deflacije eritrocita (efekt HALDANE). Tako su venski eritrociti (izravno u pluća) okrugliji od arterijskih.